
Jean-Michel Guillon
Poste : Chercheur
Poste détaillé : Chercheur CNRS
Équipe : Processus Écologiques et Pressions Anthropiques
Coordonnées :
Laboratoire Écologie, Société et Évolution – IDEEV
Bureau 2113 Bât. 680 – 12, route 128
91190 Gif Sur Yvette
Tél : +33 (0)1 69 15 57 01
Fax : +33 (0)1 69 15 73 53
Email : jean-michel.guillon (at) universite-paris-saclay.fr
Activités de recherche
La détermination environnementale du sexe chez Equisetum : mécanismes et signification évolutive.
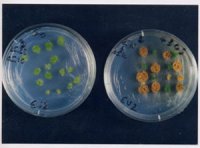
Cultures in vitro de gamétophytes de la Prêle panachée (Equisetum variegatum), montrant l’influence des conditions environnementales sur le sex-ratio. A gauche : culture sans saccharose. A droite : culture en présence de saccharose. Les gamétophytes femelles sont verts tandis que les gamétophytes mâles apparaissent oranges suite à l’accumulation de carotènoïdes.
Chez certaines espèces animales ou végétales , on observe une variation du sex-ratio (proportion de mâles) en fonction de la qualité de l’environnement. Un modèle théorique (Charnov et Bull 1977, Bull 1981) permet de prédire (i) les conditions favorisant l’apparition de ce pattern, et (ii) la valeur du biais de sex-ratio attendu. En résumé : lorsque les individus mâles et femelles ne réagissent pas de la même manière à la variation de l’environnement, l’évolution conduit à surproduire localement le sexe qui bénéficie le plus du type d’environnement rencontré. Dans ce cas, l’individu peut avoir intérêt à « choisir » son sexe en fonction de l’environnement dans lequel il se développe (détermination environnementale du sexe : ESD). L’exemple classique est celui des tortues dont le sexe est déterminé chez l’embryon par la température d’incubation.?Le modèle végétal avec lequel j’ai commencé à travailler sur ESD sont les plantes du groupe des Prêles (Equisetum).
Mon travail a d’abord consisté à décrire comment la détermination du sexe des gamétophytes pouvait être modifiée in vitro par des facteurs extérieurs comme la lumière ou la présence de sucres dans le milieu de culture (Guillon et Raquin 2002). J’ai ensuite cherché à tester la théorie de Charnov et Bull selon laquelle la lumière induirait l’apparition de femelles parce que les femelles bénéficieraient plus que les mâles d’une intensité d’éclairement élevée (Guillon et Fievet 2003). Les résultats obtenus ne soutenant pas cette hypothèse, j’ai commencé à m’intéresser à la dispersion comme facteur susceptible de modifier l’évolution du sex-ratio.
Evolution du sex-ratio et de la dispersion en environnement hétérogène.
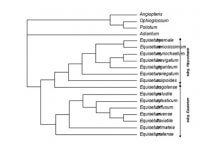
Arbre consensus obtenu par analyse Bayesienne de toutes les données de séquences ADN disponibles pour Equisetum.
L’analyse d’un modèle incorporant une dispersion biaisée selon le sexe (Guillon, Julliard et Leturque 2006) montre que des sex-ratios biaisés en fonction de l’environnement sont stables par convergence lorsqu’il existe une différence de taux de dispersion entre le pollen et les graines, ou entre les gamètes mâles et les spores chez les plantes inférieures. Cependant, les résultats de ce modèle ne permettent pas d’expliquer le pattern d’expression du sexe observé chez Equisetum. Pour mieux comprendre l’évolution de ESD chez Equisetum, j’ai reconstruit la phylogénie du genre en utilisant des gènes chloroplastiques connus pour leur transmission maternelle (Guillon and Raquin 2000). Le résultat remet en question la monoplylie des deux sous genre habituellement décrits (Guillon 2004, 2007).
Un modèle spatialisé prend en compte la dispersion d’une manière plus réaliste, en étudiant les effets de la distance de dispersion sur la stratégie de sex-ratio stable par convergence (Guillon and Bottein 2011). Un modèle étudie l’interaction entre la force sélective du modèle de Charnov et Bull, et celles qui résultent de la dispersion biaisée selon le sexe (Guillon 2016).
Une autre étude vise à comprendre les conditions requises pour l’évolution d’une dispersion biaisée selon le sexe, en introduisant une variation temporelle de la qualité de l’habitat. Nos résultats montrent que la stratégie de dispersion peut être biaisée lorsque la dispersion via les mâles se produit avant fécondation et la dispersion via les femelles après fécondation. Lorsque le taux de dispersion peut dépendre de l’habitat d’origine, les stratégies sélectionnées sont celles qui distribuent les individus dans les différents habitats selon l’espérance de la quantité de resources disponibles dans ces habitats (Aguilée, de Villemereuil et Guillon 2015).
La détermination du sexe par la température chez les reptiles : conséquences et signification évolutive.
Dans le cadre de sa thèse, Vincent Hulin a étudié l’influence de la sélection du site de ponte sur l’évolution de la détermination du sexe sensible à la tempétature (TSD) chez les tortues. En simulant une population de tortues présentant de la philopatrie natale, nous trouvons que le sex-ratio à l’équilibre est biaisé : plus de mâles sont produits là où le succès d’incubation est faible et plus de femelles sont produites là où le succès d’incubation est élevé. Ce résultat offre de nouvelles perspectives sur l’évolution de TSD chez les reptiles (Hulin et Guillon 2007). Plusieurs hypothèses pourront être testées en utilisant une nouvelle phylogénie des tortues (Guillon, Guéry, Hulin et Girondot 2012). D’autres résultats établissent une relation entre le sex-ratio des nids naturels et l’intervalle de températures de transition pour la TSD (Hulin, Delmas, Girondot, Godfrey et Guillon 2009), et évaluent les effets potentiels des changements climatiques sur les sex-ratios de tortues (Hulin, Grondot, Godfrey et Guillon 2008).
Parcours
| 2007 | HDR soutenue |
| Depuis 1998 | CR1 CNRS au Laboratoire Evolution et Systématique de l’Université Paris 11 |
| 1993 à 1997 | CR2 CNRS au Laboratoire de Biochimie de l’Ecole Polytechnique |
| 1989-1992 | Doctorat en Biologie à l’Ecole Polytechnique : Approche Enzymologique et Génétique du Rôle de la Formylation N-terminale dans le Démarrage de la Traduction chez les Bactéries |
| 1987-1988 | DEA Bases Fondamentales de l’Oncogénèse à l’Université Paris 11 |
| 1986-1987 | Agrégation de Biochimie-Génie Biologique |
| 1984-1988 | Elève à l’Ecole Normale Supérieure De Cachan (Section Biochimie) |
Publications
Girondot M, Guillon J-M (2018)-The w-value : an alternative to t- and Chi-2 tests.Journal of Biostatistics & Biometrics,1:1-3
Guillon J-M. (2007)-Molecular phylogeny of horsetails (Equisetum) including chloroplast atpB sequences. Journal of Plant Research,4:569-574 ![]()
Hulin V & Guillon J-M. (2007)-Female philopatry in a heterogeneous environment : ordinary conditions leading to extraordinary sex ratios. BMC Evolutionary Biology,7:13:1-11 ![]()
Schmitt E, Guillon J-M, Meinnel T, Mechulam Y, Dardel F & Blanquet S. (1996)-Molecular recognition governing the initiation of translation in Escherichia coli.A review. Biochimie ,78:543-554
Meinnel T, Guillon J-M, Mechulam Y & Blanquet S. (1993)-Escherichia coli fmt gene for methionyl-tRNAMetf formyltransferase escapes metabolic control. Journal of Bacteriology ,175:993-1000
Meinnel T, Guillon J-M, Mechulam Y & Blanquet S. (1993)-Structure et activité biologique: l\’exemple des acides ribonucléiques de transfert dans la traduction du message génétique en protéine. Revue Scientifique et Technique de la Défense ,20:165-172
Guillon J-M (1992)-Approche enzymologique et génétique du rôle de la formylation N-terminale dans le démarrage de la traduction chez les bactéries. Thèse de Doctorat de l’Ecole Polytechnique,134p
Guillon J-M, Mechulam Y, Schmitter J-M, Blanquet S & Fayat G. (1992)-Disruption of the gene for Met-tRNAMetf formyltransferase severely impairs the growth of Escherichia coli. Journal of Bacteriology ,174:4294-4301
Guillon J-M, Meinnel T, Mechulam Y, Lazennec C, Blanquet S & Fayat G. (1992)-Nucleotides of tRNA governing the specificity of Escherichia coli methionyl-tRNAMetf formyltransferase. Journal of Molecular Biology ,224:359-367